
Cos’è il polline?
Il polline rappresenta il gametofito maschile delle Spermatofite, cioè degli organismi vegetali più evoluti che hanno sviluppato la capacità di produrre semi (Angiosperme e Gimnosperme).
Nel ciclo vitale di tutti gli organismi con riproduzione sessuata è presente un’alternanza di fase nucleare: nella fase aploide le cellule hanno corredo cromosomico n e nella fase diploide le cellule hanno corredo cromosomico 2n.
Tale alternanza è dovuta al susseguirsi di meiosi e gamia: la meiosi produce le cellule riproduttive aploidi che si fondono nella gamia per dar vita ad un individuo diploide, che a sua volta per meiosi produrrà cellule riproduttive aploidi che fondendosi ripristinano la diploidia. Nei Metazoi (animali superiori compreso l’uomo) le uniche cellule aploidi del ciclo vitale sono i gameti e tutti gli individui sono diploidi; negli organismi vegetali, invece, all’alternanza di fase nucleare si associa l’alternanza di generazione. Nel ciclo vitale delle piante si alternano cioè 2 generazioni: lo sporofito (2n) che produce dopo la meiosi spore aploidi, ciascuna delle quali può dar vita a un gametofito (n) che produce gameti per mitosi, l’unione di due gameti porta allo sviluppo di un nuovo sporofito diploide e così via.
Angiosperme e Gimnosperme
Il polline rappresenta il gametofito maschile delle Spermatofite, cioè degli organismi vegetali più evoluti che hanno sviluppato la capacità di produrre semi (Angiosperme e Gimnosperme).
Nel ciclo vitale di tutti gli organismi con riproduzione sessuata è presente un’alternanza di fase nucleare: nella fase aploide le cellule hanno corredo cromosomico n e nella fase diploide le cellule hanno corredo cromosomico 2n.
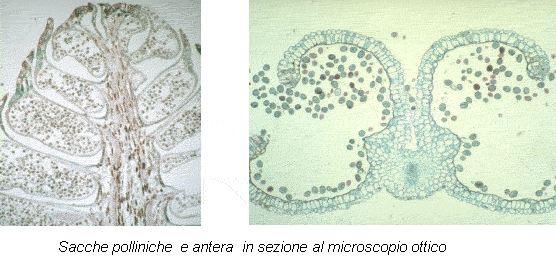
Il granulo pollinico si forma per meiosi ed è quindi una cellula aploide circondata da due pareti: una interna o intina (la cui composizione è simile a quella della parete delle cellule vegetali) ed una esterna o esina (il cui componente principale è una proteina, la sporopollenina un polimero di carotenoidi molto resistente). Per divisione mitotica il nucleo si divide in due e si ottengono un grande nucleo vegetativo, dal quale si formerà il tubetto pollinico, la struttura che consente la fecondazione, ed un nucleo generativo. Quest’ultimo si dividerà ancora una volta formando 2 nuclei spermatici che rappresentano i gameti maschili destinati a realizzare la doppia fecondazione (tipica delle Angiosperme): uno di essi feconderà il gamete femminile, la cellula uovo, dando origine all’embrione, l’altro feconderà il nucleo proendospermatico dando origine un tessuto nutritivo di riserva per l’embrione, l’endosperma secondario.
Processo di impollinazione
L’impollinazione è il trasporto del polline dall’antera allo stimma nel fiore delle Angiosperme o dalla sacca pollinica al micropilo degli ovuli nei coni femminili delle Gimnosperme. Essa può avvenire con varie modalità e, in base ai fattori che sono implicati nel trasporto del polline, si distinguono i seguenti tipi di impollinazione: anemogama, idrogama, zoogama. La prima è quella dipendente dal vento, la seconda dall’acqua e la terza dagli animali, in particolare si dice entomogama quella che avviene grazie agli insetti.

Le piante anemofile, cioè quelle che affidano al vento il compito di realizzare l’impollinazione, producono le maggiori quantità di polline, perché la loro strategia riproduttiva abbisogna di grandi quantità per avere maggiori probabilità di realizzare l’incontro casuale con l’ “obiettivo”.
Quelle entomofile invece ne producono quantità assai minori, avendo invece dei trasportatori molto efficienti e specifici.
È comprensibile quindi che le specie maggiormente rappresentate nei campioni del monitoraggio aerobiologico appartengano quasi esclusivamente a specie anemofile, sebbene molte piante si giovino di entrambi i meccanismi. Naturalmente i fiori delle piante entomofile sono quelli più vistosi e belli perché devono attirare gli insetti, mentre i fiori di quelle anemofile sono molto semplici non dovendo “far gola” a nessuno.
Il processo di impollinazione comincia quando il polline si libera dalle antere o dagli strobili per raggiungere con varie modalità i gametofiti femminili che si trovano negli ovuli, protetti da squame nei coni femminili delle Gimnosperme o dall’ovario nelle Angiosperme. Quando è giunto a destinazione il polline germina, formando il tubetto pollinico, attraverso il quale il gamete maschile raggiunge quello femminile, portando a termine la fecondazione e la formazione dello zigote diploide che diventerà l’embrione contenuto nel seme, capace di dar vita ad una nuova pianta. Il polline germina solo dopo che complessi meccanismi molecolari di riconoscimento gli confermano che è nel “posto giusto”. Questo impedisce la fecondazione tra specie diverse ma anche, per alcune piante, di fecondare gameti femminili dello stesso individuo.
Il processo di impollinazione comincia quando il polline si libera dalle antere o dagli strobili per raggiungere con varie modalità i gametofiti femminili che si trovano negli ovuli, protetti da squame nei coni femminili delle Gimnosperme o dall’ovario nelle Angiosperme. Quando è giunto a destinazione il polline germina, formando il tubetto pollinico, attraverso il quale il gamete maschile raggiunge quello femminile, portando a termine la fecondazione e la formazione dello zigote diploide che diventerà l’embrione contenuto nel seme, capace di dar vita ad una nuova pianta. Il polline germina solo dopo che complessi meccanismi molecolari di riconoscimento gli confermano che è nel “posto giusto”. Questo impedisce la fecondazione tra specie diverse ma anche, per alcune piante, di fecondare gameti femminili dello stesso individuo.granuli di polline
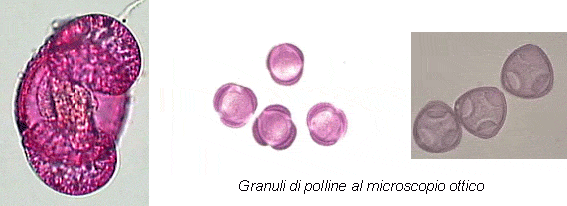
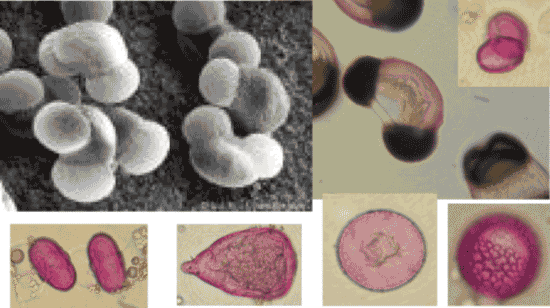
Al momento del rilascio, i granuli sono messi in libertà singolarmente, come monadi (più comuni), oppure uniti in coppia (diadi) o in altre forme di aggregazione multipla, come tetradi e poliadi. Le dimensioni variano, a seconda della specie, da un minimo di 10 μm come nelle Urticaceae ad un massimo di 200 μm (Pinaceae). Per le loro dimensioni i granuli pollinici sono visibili solo al microscopio avendo dimensioni di solito inferiori a 200 μm e generalmente comprese fra 15 e 80 μm. La disciplina scientifica che studia pollini e spore prende il nome di palinologia. Il riconoscimento palinologico è basato sull’osservazione delle caratteristiche morfologiche. Analizzando le diverse strutture al microscopio ottico, è sempre possibile determinare la famiglia di appartenenza e, per alcune entità, anche il riconoscimento a livello generico non presenta particolari difficoltà. La determinazione a livello specifico è invece possibile in pochi casi, ed in genere con l’ausilio di uno studio ecologico riferito alla zona studiata.
I pollini che più comunemente sono presenti in atmosfera sono l’espressione della flora delle diverse aree geografiche, ma soprattutto delle piante ad impollinazione anemofila, quelle cioè che liberano grandi quantità di polline durante il periodo di fioritura. Per quanto riguarda le specie ad impollinazione spiccatamente entomofila o autoimpollinanti la loro particolare biologia fiorale “non prevede” che il polline sia liberato in atmosfera. Come conseguenza il polline di queste famiglie non viene normalmente rilevato nel monitoraggio aerobiologico.
Caratteristiche strutturali e morfologiche
Il granulo pollinico è un’unità cellulare, costituita da uno strato di rivestimento e dal citoplasma, contenente il nucleo e gli organuli, e da cui trae origine il tubetto pollinico durante il processo di fecondazione dell’ovulo.
Ai fini del riconoscimento palinologico al microscopio, quello che è rilevante sono le dimensioni, la polarità, la forma e le caratteristiche della superficie esterna del granulo, in particolare la struttura della parete e le aperture. Il termine architettura della parete pollinica è stato proposto da Walcker e Doyle (1976), per includere tutti gli aspetti della morfologia della parete pollinica: la stratificazione, la struttura e le sculturazioni.
Dimensioni: i granuli di polline hanno in media dimensioni intorno ai 30 μm, ma variano dai 5 μm del Myosotis agli oltre 200 μm di alcune conifere.
Diametro maggiore di μm | Classi di dimensione | Esempi |
|---|---|---|
< 10 | Molto piccolo | Myosotis alpestre |
10 – 20 | Piccolo | Parietaria officinalis, Castanea sativa |
20 – 30 | Medio-piccolo | Quercus sp, Poa pratensis |
30 – 40 | Medio | Alnus glutinosa |
40 – 50 | Medio-grandi | Fagus sylvatica |
50 – 100 | Grandi | Zea mays |
> 100 | Molto grandi | Conifere |
Forma: in genere la forma è sferoidale, fatte salve alcune forme particolari come quella dei pollini di alcune conifere (Pinus, Cedrus, ecc.) dotati di due sacche aerifere. È il rapporto tra asse polare e asse equatoriale che definisce la forma del granulo: se le due lunghezze si equivalgono il granulo è sferico, se l’asse polare è più piccolo il granulo è oblato, se è più grande il granulo si definisce prolato.
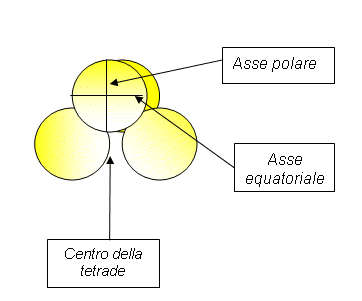
Polarità: il granulo è isopolare quando non c’è differenza tra faccia prossimale (verso il centro della tetrade) e quella distale (all’opposto); è eteropolare se le due facce sono differenti; apolare se non è riconoscibile alcuna polarità.
Struttura della parete: il citoplasma del granulo è avvolto, come tutte le cellule, dal plasmalemma e, proseguendo verso l’esterno, dall’intina e dall’esina, che vengono denominate nel loro insieme sporoderma. L’intina ha una composizione simile alla parete cellulare delle cellule vegetali, mentre l’esina è molto resistente in quanto costituita da un complesso di polimeri di carotenoidi, chiamati “sporopollenine”, che sono fra le sostanze più resistenti nel mondo organico; questo spiega la conservazione del polline fossile anche per milioni di anni. L’unico “nemico” delle sporopollenine è l’ossigeno, che ne provoca l’immediata corrosione e distruzione.
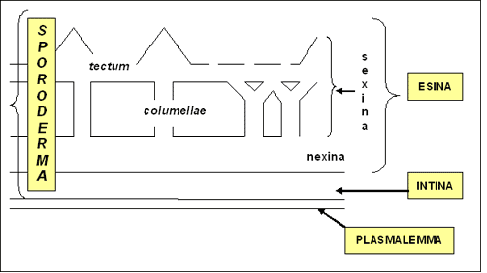

Aperture: corrispondono ai punti di uscita del tubetto pollinico e sono distinti in due tipi morfologici:
- pori di aspetto circolare o leggermente ellittico (il rapporto tra lunghezza e larghezza dell’apertura è < 2)
- colpi, lesioni allungate simili ad un solco con le estremità appuntite (il rapporto tra le due dimensioni è > 2)
A seconda delle aperture che presentano, i pollini vengono definiti come: inaperturati (se non hanno né pori né colpi), porati (quando hanno solamente pori), colpati (se presentano solo colpi) e colporati, se presentano aperture composte da un poro ed un colpo.
Anche la disposizione delle aperture può essere fondamentalmente di due tipi:
- lungo l’asse equatoriale (prefisso zono-)
- distribuiti su tutta la superficie del granulo (prefisso panto-).